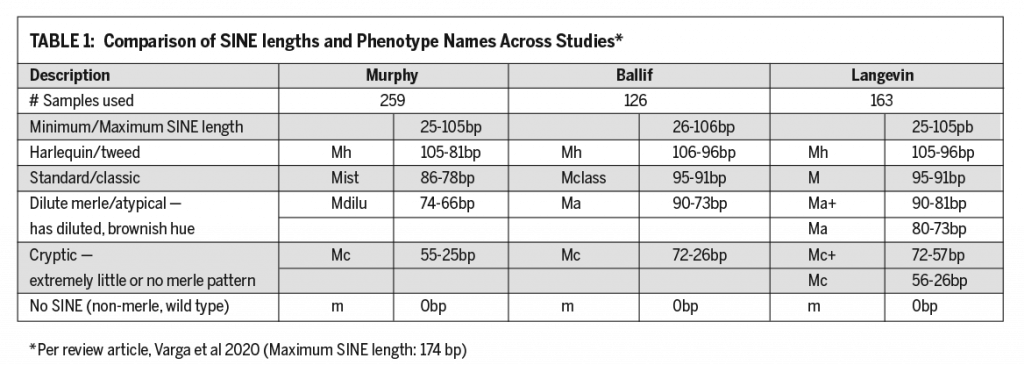
Understanding Merle and the Science Behind It.
C.A. Sharp • Grover Beach, California
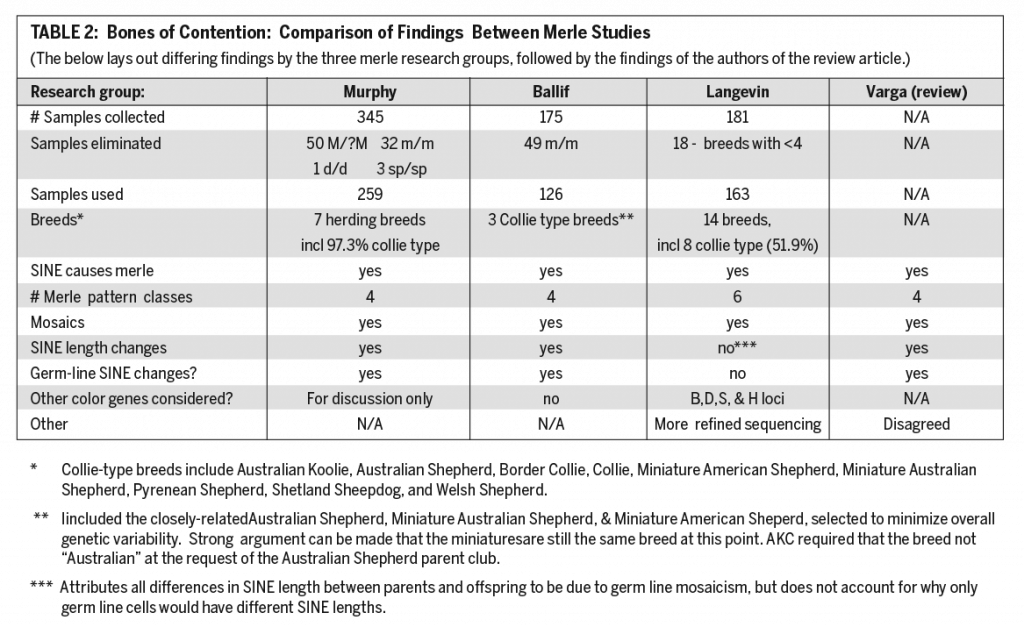
We who are involved with Australian Shepherds or other merle breeds used to think we knew all there was to be known about merle. But times change as science marches on. Tthe merle gene has been found, its structure studied, and – as sometimes happens in science – new answers lead to more questions. In the case of merle there is not total consensus amongst scientists on how to describe the pattern variations seen in merle dogs. What we do know for sure is that they are caused by a segment of DNA called a SINE (Short Interspersed Nuclear Element), the length which can vary significantly from dog to dog as well as between or within the dog’s tissues. It’s even possible for the SINE to vary from one generation to the next.
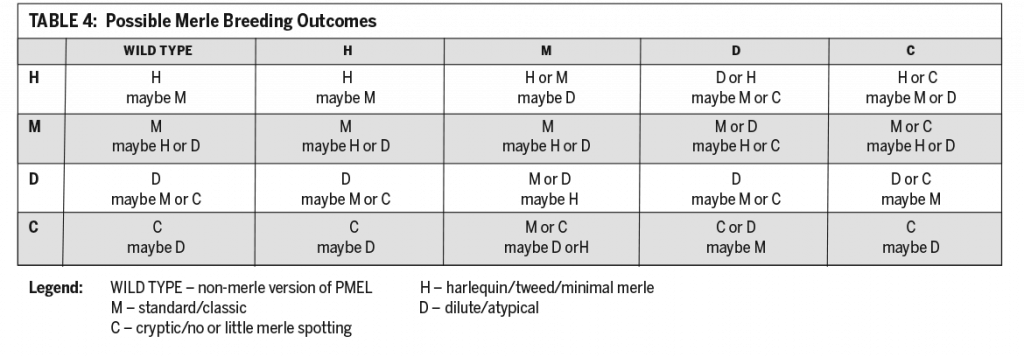
The discovery of the SINE and what it does has led to different names and symbols for variations of merle proposed by different groups of scientists. In time, further study and discussion will settle on standard descriptions and on the symbols used to represent them. In the meantime, those who breed Australian Shepherds and other merle breeds need to update their understanding of merle and how to apply that to breeding.
SINE, SINE, Everywhere a SINE…
It’s important to understand what a SINE is if you want to understand merle. Per Wikipedia, a SINE is a non-autonomous, non-coding transposable element about 100 to 700 base pairs in length. They are present in the genomes of a wide range of animal species. They likely started out as genetic parasites that moved in and settled down to stay millions of years ago. SINEs tend to be unique to a particular lineage, so a SINE you might have isn’t likely to be shared with your dog even though you are both mammals and share many of the same genes.
Since the discovery that a gene called PMEL is responsible for merle there have been three published studies that included samples from Australian Shepherds as well as another article reviewing and comparing those studies. All groups agree on the basics (which gene and how it makes dogs merle), but the version of PMEL that occurs in merle dogs is a complicated beast. The SINE can change length, causing mosaicism in the cells and tissues of the dog – and I’m not talking about the color pattern on its outside.
Gene Changes & Mosaicism
A mosaic is an individual who has different versions of the same gene in different cells of its body. Mosaicism, which can occur after conception (the union of the sperm and egg), is caused by an error in DNA replication in which a mutation is introduced or a stretch of DNA is not faithfully reproduced during cell division. This results in some cells having the exact copy of the genes inherited from the parents, while other cells carry the new mutation or stretch of DNA that was altered during replication and cell division.
The classic example of a mosaic is a calico cat. The gene that determines whether a cat is black or orange is on the female X chromosome. Therefore, if one parent was orange and the other was black, female offspring which inherit one copy of each are calico, showing a patchy arrangement of black and orange in their coats. But most mosaics, including merle dogs, may not be obvious without genetic testing.
The presence of mosaicism was one of the things the different merle investigators agreed about. A mosaic merle has more than one variant of the merle gene because of changes in SINE length after conception during cell division and gene replication. Such changes are most likely to occur during embryonic development. Descendants of a cell in which a change took place will carry that change in their DNA. The earlier it happens, the greater the number of mosaic cells the organism (in this case a dog) will have and the more types of tissue may have that new version.
Because of this, one tissue sample from a merle dog may have one length SINE while a sample from a different tissue from the same dog may have another. It is even possible that samples from the same tissue – say a liver or gonads – might include cells with different length SINES. We’ll get to details about the various studies later, but one by Langevin et al found that 6.6% of the dogs they sampled had major and minor PMEL allele variants. (For those who don’t read a lot of science, “et al” means that a study was conducted by multiple people. The first listed author plus et al is a shorthand method to avoid long lists of names. There is a list of all scientific works mentioned here at the end of the article.)
If the fact that merles can be genetic mosaics isn’t confusing enough already, just wait, there’s more! When the cell that mutated in an embryo is an ancestor of germ-line cells – the type that form the gonads – merle DNA tests may reveal a dog has a different SINE length than some or all of its offspring. And – just to confuse things a touch more – since DNA test samples are generally from buccal cells (cheek swabs) or blood, they may or may not accurately represent the SINE length present in some or all of the germ cells of that individual. If a dog’s testes or ovaries contain cells with more than one SINE length, the resulting eggs and sperm may vary in the SINE lengths they contain. When this happens the dog’s offspring may inherit different versions of merle.
To the extent that cells with differing SINE length occur in pigmented areas of the body, like the skin, irises, and hair follicles, the coloration observed can be variegated and may not correspond to what might be expected based on whatever merle variants the parents happen to have.
Another study, by Murphy et al, also found that the SINE length can change, usually becoming shorter but sometimes longer within sperm or eggs, though sperm appear to do so more often. In contrast to other investigators, Langevin et al doubt that SINE expansion occurs. They also argue that dogs with the cryptic allele, when bred to other merles, cannot produce puppies with ocular and/or hearing defects typical of double merles. They attribute this to the fact that genotyping tissues (blood and buccal cells) have different SINE lengths than those in the dog’s germ line cells and account for apparent discrepancies between genotype results and observed phenotype (appearance) in the individual dog.
There is ample research on SINES indicating that length change is generally biased toward contraction, so when a change occurs, the SINE will in most cases become shorter, rather than longer. However, SINE expansion is possible though it does not happen often. Three of the four research groups discussed here observed not only contraction but expansion among their genotyped samples.
Although most SINE insertion changes that have been observed in PMEL are contractions, many examples of SINE insertion expansions occur in other genes in genetic conditions found in humans, indicating that SINE insertion expansions in PMEL are certainly possible.
Because, the SINE almost always becomes shorter, any change in the coloration of the dog will be toward more pigment rather than less. This is why the standard/classic merle is most common and the tweed/harlequin is fairly rare. In a case where skin pigment cells and hair follicles are altered by mosaicism, you might find different merle patterns on a single dog.
Life used to be so simple!
Once upon a time there were merles who had one dominant version of the gene (M) which when coupled with the recessive version (m) gave us that patchy coat pattern we all love (M/m). Non-merles, with two copies of the recessive version (m/m), were not merle. Other genes determine whether an Aussie is red or black and bi-, tri- or without trim. Lastly, there were double merles (M/M), who had two dominant copies and – almost always in Aussies – had too much white plus potentially serious defects of the eyes and ears. Many breeders avoided this by not breeding merles to each other. But now we know it isn’t nearly so simple as that.
Merle is an endlessly fascinating, complicated, color pattern. Every merle dog seems to be is as different from any other as any given pair of human fingerprints. That’s why we who have merle breeds love it! As recent research has demonstrated, the merle gene is also amazingly complicated which is the reason there isn’t full consensus among those who have studied it closely.
It is helpful to take a look back at merle’s history – it’s origin, how we thought we had the inheritance figured out, and how we found out there was a lot more to it. I’ll finish up with a discussion of how the more recent findings apply to breeding. Since most of you reading this will be dog owners and breeders, I will do my best to use plain language as much as possible and will give a brief definition of any technical terms someone might be unfamiliar with. For those who want to delve more deeply, see that list of the studies at the end of this article.
Merle History Mystery
No one knows exactly when a gene we now call PMEL developed the mutation that causes merle coat color in dogs. The wild ancestor of all domestic dogs, the wolf, is not known for being merle. The only wild canid that has a coloration even approaching merle is the African Wild Dog (Lycaon pictus). These round-eared hunters of wild game sport a similar pattern of black blotches on a yellow background. To my knowledge, no one has ever examined the sequence of PMEL in this species. In any event, they are only distantly related to domestic dogs.
Merle may have been present in dogs for a long time, though we can’t know for sure. I once saw an ancient Egyptian painting depicting a street or market scene that included an assortment of dogs of various shapes, sizes, and colors. One was grey with black splotches. This can’t be used as proof that merle is that old – it could be nothing more than a bit of artistic license – but I did find intriguing.
The modern breeds most associated with merle largely fall into a related group we could term generic collies. Dogs of the collie type arose in Great Britain, helping farmers manage their herds and flocks. While the Rough Collie is the breed that first comes to mind when someone says “collie,” there were a wide variety of dogs of this general type all over Great Britain, living and working on farms overvarying terrain with different types of livestock. Clearly, given the frequency of merle in modern collie-type breeds, some of those early dogs must have been merle.
As Britain traded with Europe and eventually developed a global empire, British dogs, including generic collies, accompanied their people to the far reaches of the earth. Their descendants can be found in Europe, North America, Australia, and other places where the English traded or settled. Thanks to this doggie diaspora you can also find the merle pattern in a variety of breeds, some clearly not of collie type. The most notably the Dachshund and the Great Dane. These two German breeds likely obtained merle through some deep ancestral hanky-panky with a merle collie-type. In the Dane, merle is not an acceptable color but the merle version of PMEL is necessary, along with another gene, to produce the harlequin pattern of black splotches on a white background.
More recently, merle has “suddenly appeared” – most likely via a little surreptitious crossbreeding – in breeds like the French Bulldog, Pomeranian, and Chihuahua. Commercial dog breeders are similarly capitalizing on the attractive pattern’s popularity by producing a variety of -poos, -doodles, and other crossbreds (sometimes sold as a variety of pure breeds) where merle was not seen until very, very recently. The fact that the pattern is caused by the SINE insertion means it is not something likely to occur due to a spontaneous mutation.
The most important thing we know about how (though not when) merle got into dogs, is found in the gene itself.
In the beginning….
The gene called PMEL has probably been around for a very long time. It exists in a wide variety of vertebrates, including birds, reptiles, fish, and amphibians, as well as mammals. Somewhere along the way, some critter’s PMEL gene acquired a SINE. SINES are short, repetitive sequences of DNA, a type of retrotransposon, which can move around the within the genome. Where a retrotransposon chooses to relocate may affect the host gene’s function. This happened with PMEL. Retrotransposons arose from retroviral infections sometime during evolution. When a germ line (reproductive) cell is infected, the retrotransposon can be inherited by descendants of that individual. Therefore, we can thank some random ancient virus for (ultimately) giving us the lovely merle dogs we enjoy today.
PMEL is important in the development of pigment cells called melanocytes. Anything that alters PMEL may alter the coloration of the animal that inherits the new variant. Not only does it give us merle dogs, PMEL variants produce coat colors called silver in mice and horses, dominant white and smokey feather coloration in chickens, and dun in Highland and Galloway cattle, where having two variant copies makes the color lighter than in those individuals with only one. Sound familiar?
Trouble in paradise
Dog breeders have known for a long time that, however beautiful merle might be, it can also be problematic. If a dog has two merle versions of PMEL it may have eye defects, hearing loss, and more white markings than would be considered normal for a particular breed. But this isn’t true of every dog with two SINE-toting PMEL genes – some breeds are more prone than others.
The situation can be avoided easily enough: Don’t breed merles to each other. In some countries there are rules against merle/merle breeding and, as a result, the development of a merle DNA test s has complicated life for breeders in those countries, rendering dogs that would otherwise pass muster to be “unacceptable’ double merles even if they have no physical defects or disqualifying excess white.
Merle and Science
One of the earliest works pertaining to canine coat color genetics was a book: The Inheritance of Coat Color In Dogs, by Clarence C. Little DSc, a geneticist and cancer researcher who was also a breeder (Dachshunds) and show judge. He found time in his long professional career to do some work in dogs, publishing a scientific article on coat color in Great Danes in 1920, and another on the genetics of the Cocker Spaniel in 1947. Late in his career (1957) he published his coat color book. It became a standard reference for dog breeders in the era before genetic testing. The book devoted a scant four pages to merle, describing it as having only two forms, dominant M (merle) and recessive m (wild type). He also gave a brief description of the interaction between merle by the sable allele (Ay) of the A-locus (ASIP) gene.
In the 1980s, Philip Sponnenberg PhD, of Virginia Tech, published a scientific paper relating to merle. (Sponnenberg owned a blue dilute Aussie, a color arising from two recessive copies – d/d – of the D-locus gene, TYRP1.) In his article he describes what was thought to be germinal reversion – when a variant of a gene changes back into the wild type – in merle breeds, including the Australian Shepherd. He didn’t know it at the time, but he was describing a cryptic merle. Below is the abstract for that paper:
A homozygous merle Australian shepherd bitch produced two non-merle offspring and 64 that carried the merle allele. The two non-merle offspring produced no merles when bred to non-merles, indicating that the germinal reversion to non- merle was stable. This occurrence is further evidence that the merle allele is due to a transposable DNA element. [Emphasis by the author; a SINE is a “transposable DNA element.”]
In 1985, Sponnenberg and M. Lynn Lamoreaux, of Texas A&M, published the first scientific description of the tweed pattern (more recently called harlequin) in the Australian Shepherd. Tweed is a pattern with merle spots of multiple shades, sometimes including white areas on parts of the body not associated with typical Aussie white markings (a pattern seen with some frequency in the early Fairoaks line). The authors proposed that the pattern was caused by a gene they named “Tweed” (Tw). Their original proposed name for the pattern was an apt description: “Splotch.” However, during peer review they were told that the name would have to be changed because “splotch” was the name of a particular color pattern in mice. No one is apt to mix up a dog with a mouse, presumably they wanted the scientific name to be distinct and not subject to possible confusion.
What’s In a Name?
Two of the pattern names proposed for variations of merle are harlequin (also called tweed), a splotchy multi-shade pattern, and dilute merle (also called atypical merle) which features a greyish coat, sometimes with very small black spots, or in red merles, a light red base with liver spots. Each pattern is produced by a different Merle variant.
The terms dilute and harlequin are both, like Sponnenberg’s original use of “splotch,” terms that have strong association withother color genes, in this case canine color genes: D-locus (MLPH) dilute and the sort of harlequin seen in Great Danes, which have far more potential for confusion that using a term that applies to another species.
Two recessive copies of the D-locus (MLPH) gene will reduce pigment density producing lighter coat coloration: Blue dilutes would otherwise be black and “fawn’ or “Isabella” dogs would otherwise be red (also called liver). “Harlequin” is a long-standing name for a particular color pattern of black patches on a white background in the Great Dane. While merle contributes to that pattern, it requires the action of another gene, Harlequin (PSMB7) which has a variant found only in this breed that eliminates pigment in what would otherwise be blue areas leaving them white.
I would argue that the use of the terms “atypical merle” and “tweed,” (the latter has precedence by first publication: Sponnenberg and Lamoreaux 1985) should be the standard terms for these colors and would avoid confusion with other colors and patterns found in dogs caused by other genes and – in the case of PSMB7 Harlequin – one found only in a single breed
Eureka!
After that, not much happened on the merle front until the canine genome was decoded in 2005 and better gene-hunting technology developed. Since then, numerous canine genes have been identified associated with a wide variety of traits including physical features like coat type or ear carriage, various genetic diseases, and coat colors. In 2006 PMEL was identified as the source of merle (Clark et al, Proceedings of the National Academy of Science). From this point on things got much more interesting.
The initial discovery was a happy accident. Clark was studying a disease in Shetland Sheepdogs, which are sometimes merle. While trying to identify her disease gene she found a significant number of the dogs in her study had a particular form of PMEL. All of those dogs were merle. Subsequently, her group gathered and tested 259 DNA samples from dogs in seven breeds known to have merle to confirm the initial observations from the Shelties. The study was the first to note the presence of the SINE element.
Getting down to details
2018 was boom time for scientific publications focused on merle, with three separate studies on the topic.
Leigh Ann Clark, who discovered the merle gene, was a co-author on one of them (Murphy et al, Mobile DNA). Using 259 samples from 8 breeds (about a fifth were Aussies). One dog was found to be D-locus (MITF) dilute (d/d) and removed from the study as were three dogs found to be piebald (sp/sp). 85 dogs with what were described as ‘traditional’ harlequin patterns (large pigment splotches on a white background) were genotyped for the H-locus (PSMB7), but none had the dominant form that gives Danes their Harlequin pattern. Some dogs were recessive (b/b) at the B locus (TYRP1).
The group more clearly defined the SINE, which they determined to be from 25-105bp (DNA base pairs) long. They described it as being prone to “dramatic truncation” and found a clear association between differing SINE lengths and four distinct variations in merle patterning. These, from longest to shortest, were designated harlequin, standard, dilute merle, and cryptic. However, those patterns did not always fall within the expected SINE size range. They also noted that merle homozygotes (M/M) could appear to be normal merles if one allele was the short-SINE cryptic version.
Another interesting finding was that SINE length could alter during a dog’s lifetime in somatic (non-reproductive) cells, most likely during embryonic development:
Three Cardigan Welsh Corgi littermates inherited Identical Merle alleles from their sire, but display varying amounts of pigment, from typical spotting (Fig. 6a), to moderate patches (Fig. 6b), to large patches (Fig. 6c).
They further noted that the SINE sometimes changed in germ line (reproductive) cells which could result in a merle offspring having a different length variant than that of its merle parent.
Another published study (Langevin et al, PLoS One) collected 181 samples from fourteen different breeds. They excluded breeds with fewer than four individuals and used 163 samples from five breeds, all of them either collie type (50%, half of those Aussies), Catahoula (45%), and Dachshunds (5%). The largest individual breed group was the Louisiana Catahoula. (Langevin is a long-time breeder of Catahoulas which likely facilitated sample gathering among that breed community).
This group also found that SINE length could alter in non-reproductive (somatic) cells. Like Murphy’s group, they correlated specific coat patterns to SINE length, though they identified a slightly different SINE length variation (26-106bp) and described six distinct patterns, defined as harlequin, classic, atypical merle, and minimal merle, further dividing atypical and minimal merle into two sub-categories. (See Table1) They also stated that they used a more refined method of examining the SINE to better compare SINE length to phenotype.
This study was the only one to examine the effects of other color genes on merle patterning.
The third merle study (Ballif et al, Cytogenetics Genome Research) used 175 samples from three breeds. 56.6% were Aussies and 25 dogs (14.3%) were double merle. The double merles had two non-wild type versions of the gene but not always the same version.
This group also identified the presence of variants in somatic cells and suggested they may play role in color patterns which were unexpected for a given SINE length, including the small merle patches sometimes seen on cryptic merles. As with the Murphy study, they found that specific pattern did not always correspond to the expected SINE length and that cryptic paired with other variants often behaved like the wildtype version of PMEL. They also identified mosaicism in 24 (14%) of their samples, with two or more different SINE variants present in a single individual. When an offspring had a different SINE length than the parent, it was usually longer.
Regarding phenotype categories, Balif’s group noted, “…these data demonstrate that there is a continuum of merle insertion lengths associated with a spectrum of coat color and patterns and genotype-phenotype exceptions and overlap make it difficult to strictly assign certain insertion sizes with an expected coat color, although some generalizations are possible.” They also strongly rejected rigid SINE-length correlations to particular merle patterns:
…the alleles identified with various SINE insert sizes should not be interpreted as having discrete boundaries but rather should be viewed as a spectrum or continuum within which specific coat colors and patterns are likely. In addition, variations in genetic background between breeds and in individual dogs within a breed may result in the identification of different coat colors/patterns not typically found in a given size range, especially when the size of an M allele is at the border between identified groups.
Sorting it out
It is not unusual for new discoveries to have somewhat different interpretations among scientists. One way that science addresses these professional disagreements is through review articles, generally written by persons independent of the primary research. This has been the case for merle. An article published in an on-line journal in 2020 (Varga et al, Genes). [From this point forward, for the convenience of readers who don’t spend a lot of time reading research reports, the various studies will be referred to by the name of their primary author: Murphy, Langevin, Ballif , or Varga.]
Early in their article, Varga states, “Merle is among the most exciting coat color mutations of dogs.” A statement that I daresay all the various researchers involved, not to mention people in Aussies and other merle breeds, would agree. Beyond that, there are varying viewpoints sometimes difficult for lay people to understand. Hence the need for this article. Table 2 outlines the areas pf agreement and difference.
All three research groups and the reviewers agree that the SINE insertion in PMEL is the source of merle coat patterns, and that merle mosaicism occurs. Beyond that, differences arise.
Ballif and Langevin agree, with one single base pair difference, in the sign lengths related to each coat phenotype, though Langevin has subdivided two of them (see Table 1, above). Murphy assigns a somewhat longer range for harlequin/tweed/minimal merle and shorter ranges for the other three basic pattern categories. Langevin differs from the others in asserting that her subdivisions of the dilute/atypical and cryptic ranges exhibit distinct genotypes. She further holds that each base pair range within the SINE corelates to distinct coat patterns with no overlap. Murphy and Bailiff disagree with this, noting that they found each pattern sometimes occurred in dogs with a base pair range either higher or lower than that typically associated with the pattern.
What to call the various patterns also varies among the groups. Largely because Murphy and Ballif identified a different number of merle patterns (and consequently SINE lengths) than Langevin.
These nomenclature disagreements should not be surprising to dog breeders given that the name for a coloration or pattern caused by a particular gene or combination of genes in one breed can be something entirely different in another. Scientists will eventually come to some sort of consensus, dog people not necessarily. Varga’s group points out that genotype descriptive terminology was established by dog breeders in various merle breeds long before the causative gene was identified and therefore terms may vary across breeds or geographic locations, making a standard descriptive terminology acceptable to all (science, breeders, dog show judges, and specific breed communities) unlikely.
The differences in pattern categories may also stem from the differing sample sets. Murphy’s study used only collie-type breeds where half of Langevin’s samples were collie-type but half were not, those being largely Catahoula with some Dachshund. While these latter breeds do have merle, their merle patterning tends to differ from that typically seen in the collie group. This may have contributed to the differing results regarding specific merle patterns. Additionally, Langevin’s close association with the Catahoula breed may have influenced her interpretation: Of two dozen color photos in her report that are used to illustrate the different merle patterns, only five are of collie-type dogs.
The illustrations below repreent the different variations of merle, listed in order of shortest to longest SINE length. The illustrations are confined to blue merle examples without white or tan trim for simplicity and to avoid confusion. Red can vary in shade from dark chocolate through shades of brown to orange, therefore red merles can have any of these base shades in their patches on a background of dilute color. Red merle would have
liver spots where you see black and dilute red in those areas with merle patches. These illustrations are meant to provide examples of the various patterns with the understanding that the amounts of full and diluted pigment seen on any individual dog of a particular pattern can vary, sometimes widely. A dog which is not merle (wild type) will be either black or liver (red). The intensity of red pigment can also vary, so a wild type red may range from deep chocolate to medium orange or brown with merle areas lighter than the base color. Illustrations by Lisa McDonald.

Cryptic – mostly black or liver, usually with a few diluted merle spots. It is possible for a cryptic to have no visible merle.

Atypical – A mostly dilute dog with a few areas of full (black or liver) pigment. These dogs may sometimes be mistaken for the type of dilute caused by the D-locus (MLPH) gene.

Standard/Classic – Patches of black or liver on a diluted (blue or dilute red) background


Tweed/Harlequin/Herding Harlequin – these dogs have patches that vary from black or liver through a variety of diluted shades of blue or red with patches of multiple shades in the coat. These dogs sometimes have lacy areas of white in the coat as well. The term “herding harlequin” may have developed to distinguish this pattern from the genetically distinct pattern called harlequin in the Great Dane.
Murphy, Ballif, and Varga agreed that the length of the SINE can change spontaneously in both somatic and germ-line cells. Langevin argues that this occurs only in germ-line cells. In their review, Varga disagreed with Langevin that her method of determining SINE length was significantly better than that used by Murphy. They also disagreed with the assignment of two additional distinct coat pattern phenotypes, stating:
From our point of view the rigid category range system used by Langevin with the above-mentioned resolution [refers to the technology used to determine SINE length] appears unlikely, as such minimal changes in the poly(T) sequence [referring to SINE length] are not supposed to cause such drastic shifts between phenotypic categories. The overlapping category ranges in the system of Balif appear more plausible, as these authors recognized significant genotypic-phenotypic discrepancies even within
samples having the same poly(A) length… A single base-pare difference might not result in a visible phenotype category shift, and so, in our view, it would be more realistic to apply overlapping genotypic ranges in accordance with the phenotypic categories.
They further point out that Murphy had clearly established that the phenotype categories were a continuum blending one to another rather than exclusive to a particular SINE length, a finding with which Ballif agreed.
Murphy did not discuss the influence of other coat color genes where Langevin did describe interactions with double-recessive b/b and d/d genotypes.

Coat color genes can interact with each other, a prime example being the presence of both red and blue merles in the Australian Shepherd. PMEL gives the merle pattern while TYRP1 variants will determine whether it is red or blue. Murphy used other coat color genes only to exclude certain samples, likely to simplify the process of identifying patterns arising strictly from the SINE. Ballif used samples only from Aussies and Miniature American/Australian Shepherds, three very closely related groups that share the same colors and patterns. Varga suggested that in future studies the genotyping of other coat color genes along with PMEL would be advisable to determine what effects those other genes have on merle. [As an aside, in July 2021 Ballif et al. published their merle investigations in dachshunds, again concentrating their work on a single or closely related breed].
Merle and other color genes
Some other color genes can alter merle phenotype. (Consider the H-locus gene in Great Danes, which is not to be found in Australian Shepherds.) It is important for breeders to know what these are. For Aussie breeders, the most obvious and common is the recessive form of the B-locus gene (TYRP1). We like this gene because having two recessive copies means our dog will be red or red merle, both of which are acceptable per the breed standard.
Oher color gene variants are not so desirable for Aussies because they contribute to non-standard colors which the ASCA standard lists as disqualifying. The AKC standard only DQs white body splashes, but odds are if you show up in the conformation ring with a yellow dog or a blue dilute you won’t get much positive attention from the judge or your fellow exhibitors.
When you add merle into the mix there are other complications. With D-locus dilute (d/d) your blue merle will have no black spots, only shades of blue. Because of the wide variation of pigment depth in reds, it is possible a dilute red merle might not be recognized as such.
Early on there were sable (A-locus Ay) Aussies. The color was excluded in the revised ASCA standard developed in the mid-1970s. Little if any sable remains in the breed today because the Ay gene variant is dominant and we’ve been selecting away from it for nearly a half century. Given our love of merle, sable isn’t a color we’d want back – sable merles can have a muddy, unattractive coloration.
Two recessive copies of the E-locus gene (MC1r) can produce a lovely golden color, but in a merle it often evens out the pigment variation to the point that it is difficult to impossible to detect a merle pattern and therefore a good reason to exclude it. (Though they are very pretty dogs!)
If you are concerned that your dog might be or carry one of these colors, there are DNA tests available that will let you know if that is the case.
Breeding and Merle
At this point we will turn from the scientific debate to focus on what all this means for breeders of merle dogs, with the understanding that some things might shift as the scientific dust settles or new discoveries are made.
If you have a merle dog and you know that a potential mate is not merle through genetic testing (m/m), you don’t have anything to worry about. What specific pattern you will get depends on the length of the SINE the pup inherits from the merle parent, which may be shorter than that of the parent from which it was inherited. The table below shows you what you may get from a given cross. Remember that there are no longer any simple answers because there are multiple versions of merle allowing for different combinations of merle types, depending on what the parent dogs have contributed, which may differ from their own. Table 3 illustrates possible outcomes.
In the vast majority of cases, if a dog appears black or red (with or without white and/or tan), that dog will be m/m. However, it is possible that an apparently non-merle dog might have one or two cryptic versions of the gene. Usually, these dogs will have a bit of merle patterning on them somewhere, but not always. When bred, cryptic merles rarely produce what we in Aussies think of as “double merles” – the puppies will appear merle or not, though some of the “not” might also be cryptic. They aren’t likely to have hearing loss, eespecially if there is little white on the head, or merle-related eye defects. However, it can occur in about 3% of cases.
Breed may also be a factor here. Collie type breeds like the Australian Shepherd for the most part exhibit white markings of the Irish spotting pattern, featuring varying amounts of white on the face, neck, chest, under-body and legs. The gene or genes that produce this pattern have not yet been identified. The presence of the Irish pattern in collie-type breeds may amplify pigment loss in double merles. This could account for the lesser incidence of typical double merle eye and hearing issues in those merle breeds that are not of collie type, like the Catahoula.
When the SINE changes in length, getting shorter in most cases, it virtually always has a lesser impact on coat pattern, so the change is toward whatever pattern type has a shorter SINE than what you might expect based on the merle parent’s pattern.
Cryptic dogs excepted, the issue with breeding two merles together is that, statistically, a quarter of the puppies will be double merle and can have visual and hearing issues. These may be less pronounced, particularly the hearing loss, in those breeds or lines that typically have little or no white markings. For show line Aussies, where white markings are very desirable, any double merle pups are apt to have issues whereas a double merle out of a working litter from a line that carries minimal white may have no problem at all, though purposely breeding double merles is not advised.
For puppies out of a cryptic parent going into a non-breeding home, their merle status does not matter so long as they are sound and healthy. Nothing about merle will impact their health, with the possible exception of an iris coloboma (IC). This usually minor eye defect is typically seen in merle Aussies and only rarely in non-merles. At this point no one has looked at Aussies with IC who do not have a merle pattern to determine if they might have a cryptic version of PMEL. Iris coloboma can be detected in a puppy eye exam. It causes no pain and does not interfere with vision unless it is large enough to prevent the iris from contracting in bright light. A large IC will cause the dog to squint and thereby block part of the dog’s field of vision. It may also interfere somewhat with focus, but dogs do not fine-focus on tiny details the way we humans do. There is no evidence at this point that IC is caused by the PMEL gene and there are breeds that are not merle that can have this defect. IC clearly runs in families, though, so it may arise from one or more other genes in combination with the PMEL SINE.
If puppies are going to potential breeding homes and may carry the cryptic version of merle, you can either test them before they leave or advise the buyers to do so, just as you would for something like MDR1, or anything else a parent is known to carry. The result shouldn’t have any bearing on whether the pup should or should not be bred, but it should be considered in the process of mate selection.
While all this may sound daunting, all you really need to do is continue pretty much as we always have done with merle, but with the recognition that it can throw us a curve ball now and again. When in doubt, merle testing can help explain the unexpected as you make plans to avoid merle complications in the future.
The author would like to thank Lisa Schaffer PhD and Lisa McDonald MS for reviewing and offering comment on this article prior to publication.
Merle science citations
Ballif, BC; Ramirez, CJ; Carl, CR; Sundin, K, Krug, M; Zahand, A; Shaffer, LG; Flores-Smith, H “The PMEL Gene and Merle in the Domestic Dog: A continuum of Insertion Lengths Leads to a Spectrum of Coat Color Variations in Australian Shepherds and Related Breeds” Cytogenetics Genome Research 2018 156, 22-34
Bailif, BC; Ramirez CJ; Carl, CR; Sundin, K; Flores-Smith, H; Schaffer, LG “The PMEL gene and merle (dapple) in dachshunds: cryptic, hidden, and mosaic variants demonstrate the need for genetic testing prior to breeding” Human Genetics https://doi.org/10.1007/s00439-021-02330-y
Clark, LA; Wahl, JM; Rees, CA; Murphy, KE “Retrotransposon Insertion in SILV is responsible for merle patterning in the domestic dog” Procedures of the National Association of Science USA 2006 103, 1376-1381
Langevin M; Symkova, H; Jancuskova, T, Pekova, S “Merle Phenotypes in Dogs-SILV SINE Insertions From Mc to Mh” PLOS One 2018 13, e0198536.
Little, Clarence C., The Inheritance of Coat Color in Dogs, p 70-73, 1957. Original publisher Comstock Publishing Assoc. and later Howell Book House.
Murphy, SC; Evans, JM; Tsai, KI; Clark, LA “Length variations with in the Merle retrotransposon of canine PMEL: Correlating genotype with phenotype” Mob DNA 2018,9, 26.
Sponnenberg, P “Germinal reversion of the merle allele in Australian shepherd dogs” Journal of heredity 1984 p78
Sponnenberg, P; Lamoreaux, ML “Inheritance of tweed, a modification of merle, in Australian Shepherd Dogs” Journal of Heredity, July 1985, p.303-4
Varga, L; Lenart, X; Zenke, P; SLazlo, O; Hudak, P, Ninausz, N, Pelles, Z, Szoke, A “Being Merle: The Molecular Genetic Background of the Canine Merle Gene” Genes 2020, 660,doi: 10.3390/genes11060660